- Date :21 Jan, 2021
- カテゴリー : 物理化学
岡山大学異分野基礎科学研究所の墨智成准教授および豊橋技術科学大学情報・知能工学系の原田耕治助教は、海馬興奮性ニューロンにおける長期増強(LTP)と長期抑制(LTD) の発現を統一的に説明する分子機構を提案し、大規模数理モデルシミュレーションを用いて実証しました。
これまでLTPとLTDはそれぞれ個別に説明が試みられてきましたが、それらを統一的に説明する試みはなされていませんでした。本研究では、海馬興奮性ニューロンの後シナプスにおけるAMPA型グルタミン酸受容体(AMPAR)の「能動的」輸送過程を正確に再現した大規模数理モデルを提案し、それに基づくシミュレーション実験を行いました。その結果、NMDA型グルタミン酸受容体を介して後シナプスに流入するカルシウムイオン濃度の多寡に依存した「シナプトタグミン1/7によるエキソサイトーシス」および「PICK1によるエンドサイトーシス」の競合が、シナプス後膜上のAMPAR数を増減させ、その結果としてLTPおよびLTDが誘導されることを明らかにしました。本研究成果は英国の科学雑誌「Scientific Reports」に掲載されました。
本研究は、LTPおよびLTDへの関与が特定されているタンパク質(並びに複合体)の、AMPAR輸送システムにおける役割や、1分子計測によって観測されているさまざまな側面を、包括的に理解・説明するための分子基盤となることが期待されます。
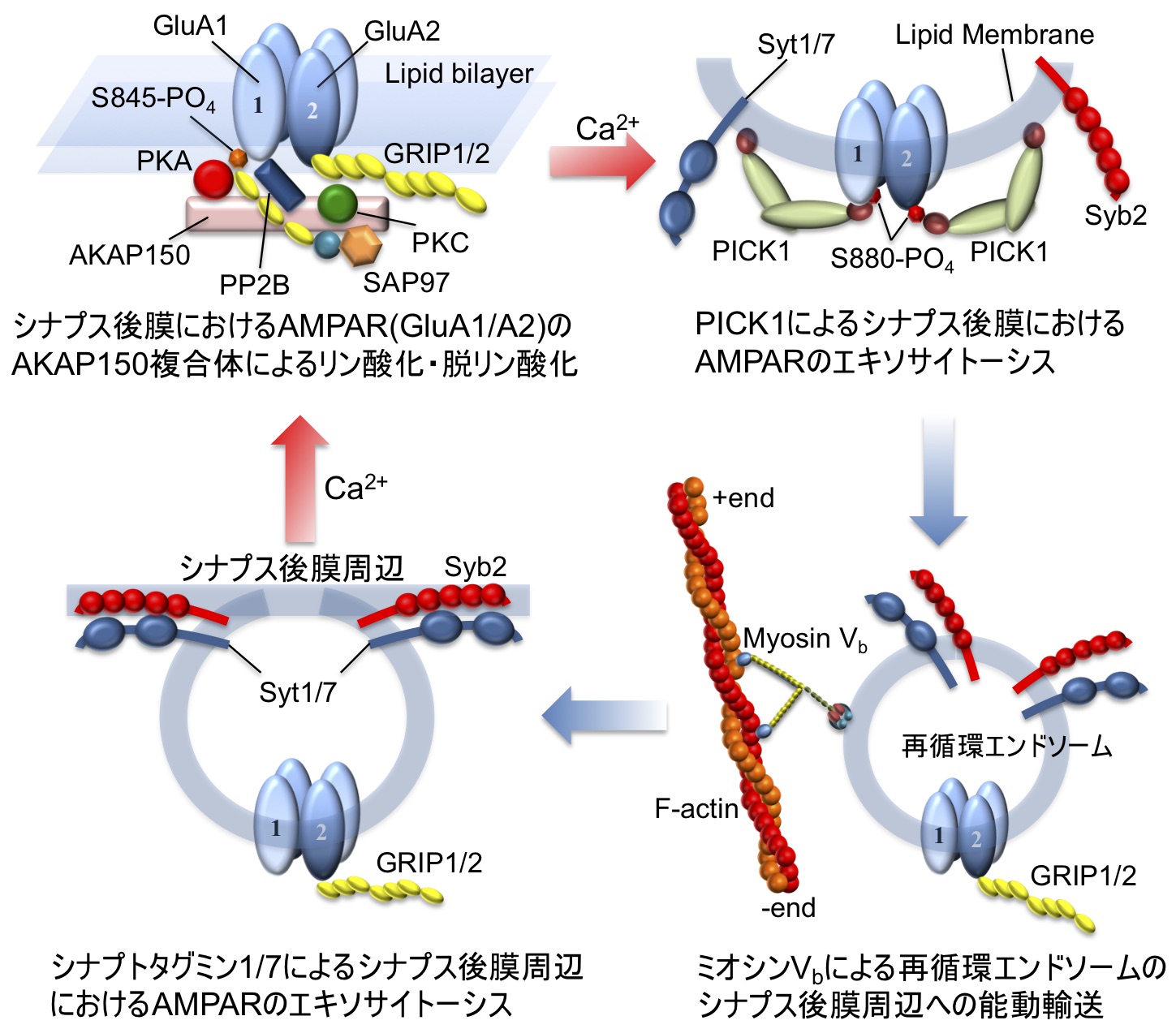
図1. 海馬興奮性ニューロンにおけるLTP/LTDを再現する後シナプスにおけるAMPAR輸送システム。(左上) AKAP150シグナル複合体(リン酸化酵素PKA, PKC; 脱リン酸化酵素PP2B)と GluA1/A2ヘテロ4量体)AMPARとの間の足場タンパク質(SAP97, GRIP1/2)を介した相互作用モデル。(右上) カルシウムセンサPICK1によるAMPARのカルシウム依存エンドサイトーシス。(右下) エンドサイトーシスで細胞質内に取り込まれたAMPAR含有再循環エンドソームの分子モーターミオシンVbによるシナプス後膜周辺方向への能動的輸送。(左下) カルシウムセンサシナプトタグミン1/7(Syt1/7)によるAMPAR含有再循環エンドソームのカルシウム依存エキソサイトーシスによるシナプス後膜周辺へのAMPARの取り込み。
論文情報
-
論文名:Mechanism underlying hippocampal long-term potentiation and depression based on competition between endocytosis and exocytosis of AMPA receptors
-
掲載誌:Scientific Reports
-
著者:Tomonari Sumi, Kouji Harada
なお、本研究の記事は、科学新聞で紹介されました。

- 科学新聞2020年10月2日掲載
- 科学新聞社の許可を得て掲載しております。転載はご遠慮ください。